
Введение
С момента внедрения способов получения рекомбинантной ДНК в течение последних двадцати лет постоянно увеличивается количество поступающих на рынок биологических лекарственных препаратов, таких как инсулин, гормон роста человека, интерфероны и эритропоэтин. Истечение срока действия патентов на первое поколение биофармацевтических препаратов способствует разработке и регистрации т. н. биоаналогов, или воспроизведенных биологических лекарственных препаратов. В отличие от более распространенных низкомолекулярных лекарственных препаратов, биологические препараты обычно имеют высокомолекулярную структуру и могут быть довольно чувствительными к изменениям процесса производства. Производитель воспроизведенного биологического лекарственного препарата не имеет доступа к молекулярному клону оригинального препарата и оригинальному банку клеток, а также не имеет возможности в точности воспроизвести процессы ферментации и очистки. Даже использование тех же клеточных линий и генов, аналогичных процессов производства, очистки и создания готовых лекарственных форм не гарантирует полную эквивалентность воспроизведенного (биоаналога) и инновационного биологического препарата.
Это означает, что два аналогичных биологических лекарственных препарата могут вызывать различный иммунный ответ. Поэтому, в отличие от химически синтезированных фармацевтических препаратов, замещение биологических препаратов биоаналогами, может иметь клинические последствия и представлять определенную опасность для здоровья.
Одним из наиболее продаваемых биологических лекарственных препаратов является эритропоэтин. Он представляет собой гликопротеиновый гормон, который главным образом вырабатывается в почках и стимулирует образование эритроцитов в костном мозге [1]. Рекомбинантный эритропоэтин человека, полученный методом генной инженерии, успешно применяется для лечения анемии, развивающейся на фоне хронической почечной недостаточности [2].
Молекулярная масса эритропоэтина составляет 30–34 кДа, причем 40 % молекулярной массы обусловлено углеводородами, из которых 17 % приходится на сиаловую кислоту [3–6]. Эритропоэтин – гетерогенный протеин, состоящий из нескольких различных изоформ, образующихся преимущественно путем гликозилирования [7]. Сахара присоединяются к протеину в участках гликозилирования путем образования одной O-связи (серин) и трех N-связей (аспарагин). Гликозилирование имеет большое значение для биологической активности и стабильности эпоэтина [8–12].
Производством эпоэтина занимается несколько компаний. В России рекомбинантный эритропоэтин человека продается под торговыми названиями Эпрекс* и Эральфон*. Результаты сравнительного исследования применения нескольких эпоэтинов, проведенного H. Schellekens (2002), показали, что препараты статистически значимо различаются по биологической активности и изоформному составу [13]. Автор пришел к выводу о том, что качество проанализированных биоаналогов вызывает сомнения. В исследование H. Schellekens (2002) не вошли препараты, продаваемые в России.
В нашей работе два доступных в России препарата эпоэтина, Эпрекс* и Эральфон*, были сопоставлены между собой с использованием двумерного гель-электрофореза и электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (ДСН-ПААГ электрофореза) с вестерн-блотом.
Материалы и методы исследования
2.1. ОБРАЗЦЫ
Образцы были приобретены в аптеках и доставлены в исследовательскую лабораторию в контролируемых условиях при температуре 2–8 °C. Сведения о названии каждого препарата, производителе, стране и номере серии представлены в табл. 1.
2.2. ОБОРУДОВАНИЕ И ПРОЦЕДУРЫ
2.2.1. УДАЛЕНИЕ АЛЬБУМИНА
Удаление альбумина было выполнено в соответствии с протоколом набора для удаления ProteoPrep20 компании “Sigma” (prot20). Емкость колонки ProteoPrep20 составляет 300 мкг альбумина; соответственно, для удаления примерно 1 мг альбумина из каждого шприца необходимо четыре раза подряд пропускать через колонку. По 2 шприца каждого из исследуемых препаратов (Эпрекс* и Эральфон*) концентрировали с помощью Центрикон (5 кДа, “Millipore”), промывали 1,5 мл уравновешивающего буфера, после чего доводили объем до 800 мкл с помощью уравновешивающего буфера. В уравновешенную колонку вносили 100 мкл промытого продукта, инкубировали в течение 20 минут и затем центрифугировали (1 минута, 2000 оборотов/мин). Далее промывали колонку 2 раза с помощью 100 мкл уравновешивающего буфера. Все фракции собирали и концентрировали Центриконом (5 кДа, “Millipore”) до 40 мкл.
2.2.2. ПРИГОТОВЛЕНИЕ ОБРАЗЦОВ ДЛЯ ДВУМЕРНОГО ГЕЛЬ-ЭЛЕКТРОФОРЕЗА
Во время размораживания образцов добавляли мочевину, амфолиты и дитиотреитол (ДТТ) таким образом, чтобы конечная концентрация составила 9 M мочевины, 2 % амфолитов и 70 мM ДТТ. После инкубации в течение 30 минут при комнатной температуре и центрифугирования в течение 45 минут со скоростью 15 тыс. оборотов/мин супернатант отбирали и замораживали в новых пробирках при температуре -80°C.
2.2.3. ДСН-ПААГ ЭЛЕКТРОФОРЕЗ
ДСН-ПААГ электрофорез был проведен в соответствии с методикой, описанной Laemmli и соавт. [11]. По 2 мкл концентрированного образца без добавления мочевины, с добавлением мочевины и без добавления ДТТ и с добавлением мочевины и ДТТ наносили на каждую дорожку ДСН-ПААГ (15 %-ный акриламид (30 : 1); 0,1 %-ный додецилсульфат натрия; 0,06 %-ный тетраметилэтилендиамин; 0,02 %-ный аммония персульфат), после чего проводили электрофорез в течение 75 минут при 150 В до тех пор, пока фронт не достигал кромки геля. После разделения гели окрашивали красителем FireSiver (“Proteome Factory”, PS-2001) или подвергали блоту.
2.2.4. ДВУМЕРНЫЙ ГЕЛЬ-ЭЛЕКТРОФОРЕЗ
По 20 мкл очищенного и концентрированного протеина наносили на гели для вертикального электрофореза (9 M мочевины, 4 %-ный акриламид; 0,3 %-ный картофельно-декстрозный агар, 5,0 %-ный глицерол; 0,06 %-ный тетраметилэтилендиамин и 2,0 %-ный носитель амфолитов (pH 2–11); 0,02 %-ный аммония персульфат) для изоэлектрического фокусирования при 8820 В-ч в первом направлении. После изоэлектрического фокусирования гели инкубировали в уравновешивающем буфере, содержащем 125 мM трисфосфата (pH 6,8), 40 % глицерола, 65 мM ДТТ и 3 % ДСН, в течение 10 минут, и затем замораживали при температуре -80°C. Готовили гели для проведения ДСН-ПААГ электрофореза во втором направлении (8 см x 9 см x 0,15 см), содержащие 375 мM буфера трис-HCl (pH 8,8), 15 % акриламида, 0,2 % бисакриламида, 0,1 % ДСН и 0,03 % тетраметилэтилендиамина. После размораживания уравновешенные гели (после изоэлектрического фокусирования) немедленно наносились на гели для ДСН-ПААГ-электрофореза. Электрофорез проводили в течение 75 минут при 150 В до тех пор, пока фронт не достигал кромки геля. После двумерного разделения гели окрашивали красителем FireSilver (“Proteome Factory”, PS-2001).
2.2.5 ВЕСТЕРН-БЛОТ
Вестерн-блот проводили при следующих условиях (табл. 2):
2.2.6. ИММУННОЕ ОКРАШИВАНИЕ
Способ иммунного окрашивания приведен ниже.
1. Промыть 2 раза в TBST (20 мM трис, pH 7,5; 154 мM NaCl, 0,1 % Tween 20).
2. Блокировать 2 %-ным раствором бычьего сывороточного альбумина в TBST в течение 2 часов.
3. Инкубировать с первичными антителами (против эритропоэтина, “Sigma”, E2531) в разведении 1 : 1000 в 1 %-ном растворе бычьего сывороточного альбумина в TBST в течение ночи.
4. Промыть 5 раз по 10 минут в TBST.
5. Инкубировать с вторичными антителами (антитела к IgG кролика, конъюгированные с щелочной фосфатазой, “Sigma”, A3812) в разведении 1 : 1000 в 1 %-ном растворе бычьего сывороточного альбумина в TBST в течение 2 часов.
6. Промыть 5 раз по 10 минут в TBST.
7. Проявить с помощью Fast Red/Naphtol (“Sigma”) в течение 5 минут.
8. Остановить реакцию с помощью воды.
Результаты
Препарат Эральфон* содержит существенное количество человеческого альбумина, который может препятствовать разделению белков. Поэтому альбумин был удален с помощью колонки. Для обеспечения надежности результатов через колонку были пропущены образцы обоих препаратов Эпрекс* и Эральфон*.
3.1. АНАЛИЗ РЕЗУЛЬТАТОВ ОДНОМЕРНОГО ВЕСТЕРН-БЛОТА
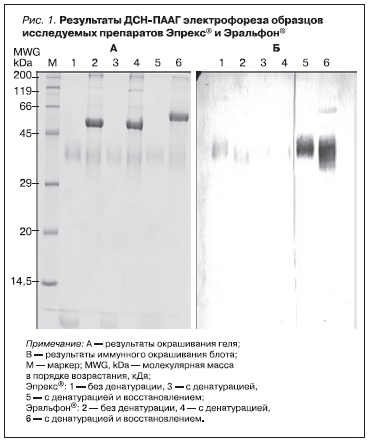
После удаления альбумина и концентрации образцы исследуемых препаратов Эпрекс* и Эральфон* объемом 2 мкл были разделены путем 15 % ДСН-ПААГ электрофореза один раз без денатурации (без добавления мочевины), один раз с денатурацией, но без восстановления (с добавлением мочевины и без добавления ДТТ) и один раз с денатурацией восстановлением. Часть гелей была сразу же окрашена, другая часть была перенесена на мембрану из поливинилденфторида и подвергнута блоту. После проведения блота было выполнено иммунное окрашивание мембраны из поливинилденфторида. В качестве первичных антител использовались антитела против эритропоэтина в разведении 1 : 1000. В качестве вторичных антител использовались антитела к IgG кролика, конъюгированные со щелочной фосфатазой, в разведении 1 : 1000. Первые полоски проявились через 30 секунд; через 5 минут реакция была остановлена.
На геле на дорожках 1–6 (рис. 1A) определяются слабые полоски между 32 и 44 кДа. В образце препарата Эпрекс* больше никаких полосок не определяется. В образце препарата Эральфон* определяются дополнительные полоски: четкая полоска около 50 кДа на дорожке 2 и 4 и около 60 кДа на дорожке 6, а также слабая полоска на уровне 200 кДа и еще одна полоска на уровне 120 кДа на дорожке 4. Результаты иммуноблотинга (рис. 1B) подтвердили, что слабые полоски на рис. 1A между 32 и 44 кДа соответствуют эритропоэтину. Молекулярные массы эритропоэтина в препарате Эпрекс* находятся в узком диапазоне 40–45 кДа, тогда как молекулярные массы эритропоэтина в препарате Эральфон* находятся в диапазоне 32–45 кДа. Это свидетельствует о более высокой степени гликозилирования эритропоэтина в препарате Эпрекс* по сравнению с эпоэтином в препарате Эральфон*. Надлежащее гликозилирование может играть важную роль в биологической активности. Другие полоски геля не вступают в реакцию с антителами против эритропоэтина, что указывает на то, что они образованы не эритропоэтином. Агрегатов эритропоэтина не выявлено. Таким образом, были выявлены явные различия между препаратами Эпрекс* и Эральфон*.
3.2. АНАЛИЗ РЕЗУЛЬТАТОВ ДВУМЕРНОГО ВЕСТЕРН-БЛОТА
Был проведен двумерный гель-электрофорез образцов исследуемых препаратов Эпрекс* и Эральфон* (рис. 2, 3). По одному гелю каждого типа было окрашено красителем FireSilver, второй гель был подвергнут блоту и иммунному окрашиванию. После двумерного электрофореза образца препарата Эпрекс* на геле определяются три пятна в области изоэлектрического фокусирования для pH 3–4. Больше никаких пятен не выявлено, в отличие от препарата Эральфон*, на геле которого обнаружено много пятен. Результаты иммунного окрашивания блота после двумерного гель-электрофореза образца препарата Эральфон* свидетельствуют о положительной реакции для 7 пятен в области изоэлектрического фокусирования для pH 3–5. Другие пятна, выявленные на окрашенном FireSilver геле после двумерного электрофореза, не взаимодействуют с антителами против эритропоэтина. Они могут быть образованы альбумином, не полностью удаленным при пропускании через колонку.
Таким образом, препарат Эральфон* содержит существенно большее количество изоформ эритропоэтина по сравнению с препаратом Эпрекс*. При исследовании обоих препаратов агрегатов не выявлено.
Результаты нескольких исследований свидетельствуют о том, что различные изоформы эпоэтина обладают различной биологической активностью [6–9]. Поэтому биологическая активность препаратов Эпрекс* и Эральфон* может различаться. В препарате Эральфон* содержится несколько изоформ, которые являются более основными, чем изоформы, входящие в состав препарата Эпрекс*. Это может свидетельствовать о меньшем периоде полувыведения эпоэтина [8, 12].
Выводы
В ходе исследования выявлены различия между образцами двух анализируемых препаратов Эпрекс* и Эральфон*. Полученные данные свидетельствуют о том, что препарат Эральфон* содержит больше изоформ эритропоэтина, образовавшихся в результате неполного гликозилирования, и они имеют более щелочные характеристики, что может свидетельствовать о более коротком периоде полувыведения Эральфона*. Выявленные различия степени гликозилирования могут вызвать нежелательный иммунный ответ, что может привести к неблагоприятным клиническим последствиям [13]. По данным международных исследований биологические активности разных изоформ эпоэтина различны [6–9]. Поэтому биологическая активность препаратов Эпрекс* и Эральфон* может отличаться.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}